Vision in detail
$0.00
This article explores vision from the human eyeball to the primary visual cortex in detail (and one step at a time).
The article is in PDF format and available for free download.
Description
This article will soon be available for free download. Select PDF in the dialogue box above and add to downloads.
Vision in detail
AN INTRODUCTION TO COLOUR VISION
1.1 - Human eye in cross-section

1.2 - Experience of seeing
Experience of seeing
Colour is something we see every moment of our lives if we are conscious and exposed to light. Some people have limited colour vision and so rely more heavily on other senses – touch, hearing, taste and smell.
Colour is always there whether we are aware and pay attention to it or not. Colour is what human beings experience in the presence of light. It is important to be clear about this. Unless light strikes something, whether it is air, a substance like water, a physical object or the retina at the back of our eyes, light, as it travels through space, is invisible and so has no colour whatsoever. colour is an artefact of human vision, something that only exists for living things like ourselves. Seeing is a sensation that allows us to be aware of light and takes the form of colour.
The experience of colour is unmediated. This means that it is simply what we see and how the world appears. In normal circumstances, we feel little or nothing of what is going on as light enters our eyes. We have no awareness whatsoever of the chemical processes going on within photosensitive neurons or of electrical signals beginning their journey towards the brain. We know nothing of what goes on within our visual cortex when we register a yellow ball or a red house. The reality is, we rarely even notice when the world disappears as we blink! In terms of immediate present perception, colour is simply something that is here and now, it is that aspect of the world we see as life unfolds before us and is augmented by our other senses, as well as by words, thoughts and feelings etc.
It takes about 0.15 seconds from the moment light enters the human eye to conscious recognition of basic objects. What happens during this time is related to the visual pathway that can be traced from the inner surface of the eyeball to the brain and then into our conscious experience. The route is formed from cellular tissue including chains of neurons some of which are photosensitive, with others tuned to fulfil related functions.
So, let’s start at the beginning!
Before light enters the eye and stimulates the visual system of a human observer it is often reflected off the surfaces of objects within our field of view. When this happens, unless the surface is mirror-like, it scatters in all directions and only a small proportion travels directly towards our eyes. Some of the scattered light may illuminate the body or face of an observer or miss them completely. Some is reflected back off the iris enabling others to see the colour of their eyes. Sometimes light is also reflected off the inside of our eyeballs – think of red-eye in flash photography.
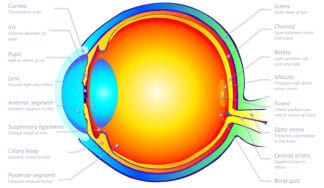
 Cross-section of the human eyeball
Cross-section of the human eyeball
The fraction of light that really counts passes straight through the pupil and lens and strikes the retina at the back of the eyeball. From the point of view of an observer, this leads to two experiences:
- Things an observer sees right in the centre of their field of vision, which is to say, whatever they are looking at.
- Things an observer sees in their peripheral vision and so fill the remainder of a scene.
If we think of light in terms of rays, then the centre of the field of vision is formed from rays that enter our eyes perpendicular to the curvature of the cornea, pass right through the centre of the pupil and lens and then continue in a straight line through the vitreous humour until they strike the retina. Because these rays are perfectly aligned with our eyeballs they do not bend as they pass through the lens and so form an axis around which everything else is arranged. The point where this axis strikes the retina is called the macula and at its centre is the fovea centralis where the resulting image appears at its sharpest.
Peripheral vision is formed from rays that are not directly aligned with the central axis of the cornea and pupil, and do not pass through the very centre of the lens. All the rays of light around this central axis of vision change direction slightly because of refraction.
It deserves mention at this point that the lenses in each eye focus in unison to accommodate the fact those things we scrutinise most carefully may be anywhere from right in front of our noses to distant horizons.
We must also not forget that the optical properties of our lenses mean that the image that forms on the retina is both upside down and the wrong way round.
1.3 - Visual processing
Visual processing
Visual processing is a complex and dynamic process that involves interactions between various retinal cells, neural pathways, and brain regions, ultimately leading to conscious visual perception.
Visual processing begins the moment light enters the human eye. It then progresses through multiple stages as signals travel towards the visual cortex, where the neural activity is integrated, resulting in conscious visual experience.
As visual processing begins the retina starts to process information about colors, as well as basic information about the shape and movement associated with those colors. By the end of this stage, multiple forms of information about a visual scene are ready to be conveyed to higher brain regions.
Let’s examine two major forms of processing, trichromatic and opponent-processing, which occur within the eyeball as visual information is gathered from light entering our eyes.
Trichromacy, also known as the trichromatic theory of colour vision, explains how three types of cone receptors in the retina work together with bipolar cells to perform their role in the initial stage of colour processing. Rod cells also play a significant role in this form of processing visual information, particularly in low-light conditions.
Opponent-processing, also known as the opponent-process theory of colour vision, explains the second form of processing. Opponent-processing involves ganglion cells that process the data received from trichromatic processing and combine it with other intercellular activities.
It is interesting to note that as both trichromatic and opponent-process theories developed over the last century, researchers and authors have often pitted one theory against the other. However, both processes are crucial for understanding how colour vision occurs.
Trichromatic theory explains the encoding of visual information when light hits the retina, while opponent-processing explains a subsequent stage of information convergence, assembly, and coding before the data leaves the retina via the optic nerve.
Note that:
- Both trichromatic and opponent-processing occur independently within each retina, without comparing with the other.
- Each eye gathers information from a specific viewpoint, approximately 50 mm to the left or right of the nose.
- The two impressions are later compared and combined to provide us with a single three-dimensional, stereoscopic view of the world, rather than two flattened images.
We can consider the layers of retinal cells involved in trichromatic and opponent-processing as examining, interpreting, and transmitting visually relevant information. However, it would be incorrect to view this as a straightforward linear process due to the intricate neural networking, cross-referencing, and feedback loops within the retina.
1.4 - Trichromatic processing
Trichromatic processing
To get some sense of what is going on during the trichromatic processing of visual information collected during phototransduction within rod and cone cells, we need to imagine the light-receiving layer within the retina of each eye as being composed of around 120 million points. Each point is the location of a rod or cone within a mosaic of cells stretching to the boundary of the retina in every direction. Each point also corresponds with a different location within the overall image of the world outside that is projected onto the retina by the lens as light enters the eyes through the cornea, and pupil.
Every rod and cone cell within the human eye is an independent photo-sensitive neuron that responds independently to incoming photons of light. In the first instance, each one carries out its task of responding to the stream of different wavelengths of light without reference to the receptors around it. We must imagine all these neurons as being capable of functioning simultaneously and being able to fire several times a second. Each time a cell responds, phototransduction takes place, which is a chemical response to light that produces electrical impulses ready for transmission.
What precisely happens depends on lighting conditions.
When light levels are low, each photosensitive rod cell may be hit by a handful of photons of a given wavelength and in some cases, this is enough to trigger the chemical response. But at very low levels of lighting, the response of cone cells is very limited. The outcome is a typical night scene that appears blue-purple-black with very little detail.
When light levels are high, each cone cell may be bombarded with tens of thousands of photons of a given wavelength within the briefest fraction of a second, producing a powerful chemical response. In these conditions, rod cells tend to be overwhelmed by the sheer quantity of light but can still provide useful information in peripheral vision. The outcome is a world rich in colour, full of detail and with contrasting highlight-detail and deep well-defined areas of shadow.
The trichromatic process involves the sampling of this mosaic of rod and cone photoreceptors by bipolar and horizontal cells. Individual photoreceptors are connected in small groups to bipolar cells that receive their electrical output. The bipolar cells compare the response of receptors with the response of neighbours, whilst horizontal cells help to aggregate the result and encode it into three independent data channels.
The comparison process involves what is termed centre-surround antagonism. This refers to the way bipolar neurons organize their receptive fields. A close-up view reveals that centre-surround antagonism relies on small groups of cells being arranged around a centre point where one rod or cone synapses on the dendrites of a bipolar cell. Around this centre are other photoreceptors that also synapse onto other dendrites of the same bipolar cell. The signal received from the centre is compared with a summation of the signals from neighbours to establish to what extent they agree or disagree. This process goes on in real-time as bipolar cells receive successions of signals and horizontal cells modulate the information to improve fidelity. The scale of this enterprise as it takes place across the surfaces of the retina in each eye and in real-time is extraordinary.
The role of horizontal cells in both trichromatic and centre-surround antagonism is best conceived as signal conditioning and particularly with globally adjusting visual information ready for opponent processing. However, ongoing research suggests that horizontal cells may also be involved in an early stage of signal interpretation and so contribute towards things such as the detection of edges and movement.
Trichromatic colour vision is governed by the fact that there are three distinct types of cone cells within the retina, each tuned to respond to a different band of wavelengths of light. As light floods in, each type of cone outputs a signal if it picks up the presence of photons with wavelengths it recognises. Every identifiable point on the retina contains all three types of cone cells and these are tightly packed into a random mosaic pattern across the entire surface. The result is a photosensitive film containing millions of receptors capable of responding to all wavelengths across the visible spectrum.
General descriptions of trichromatic vision often suggest that the three types of cone cells in the human retina are responsive to wavelengths corresponding to red, green and blue. It is more accurate to say that the peak sensitivity of these L, M and S cones types respond with biases towards different regions of the visible spectrum and have a loose correspondence with red, green and blue:
- L cones: Respond to long wavelengths (peak sensitivity around 560 – 580 nanometres) so with a region of sensitivity that includes red, orange, green and yellow but with a peak bias between red and yellow.
- M cones: Respond to medium wavelengths (peak sensitivity around 530 – 545 nanometres) so with a region of sensitivity that includes orange, green, yellow and cyan but with a peak bias between yellow and green.
- S cones: Respond to short wavelengths (peak sensitivity around 420 – 440 nanometres) so with a region of sensitivity that includes cyan, blue and violet but with a peak bias between blue and violet.
- Rods: Rod cells are most sensitive to wavelengths around 498 nanometres, so with a peak sensitivity towards green-blue, and are insensitive to wavelengths longer than about 640 nanometres (red).
This arrangement suggests that:
- At any specific moment, all three types of cone cells at any specific location on the retina may fire multiple times per second in response to streams of photons that have constantly changing wavelengths.
- The assessment of the distribution of wavelengths by every individual cone depends on both its range of sensitivity and the wavelength at which its response peaks.
- Centre-surround comparisons check for consistencies and variations within the responses of cone groupings at every location.
- As soon as a clean and noise-free consensus is achieved, the data specific to each cone type is output on one of three separate data channels.
- Once the composite of trichromatic and centre-surround encoding of colour information is complete it is sent onward for opponent processing within the retina.
- Recent research suggests that the three output channels do not correspond directly with the L, M and S cone types. There are indications that two channels contain chromatic information whilst the other contains achromatic data.
1.5 - Opponent-processing
Opponent-processing
The trichromatic and centre-surround processing described above creates a time-coded, high fidelity, redundancy-free stream of information ready to be passed forward to ganglion cells where opponent-processing takes place and colour information is even more rigorously and economically encoded.
Opponent-processing is believed to represent a shift in emphasis from the physiologically orientated mechanisms of trichromatic processing towards data-types more adapted to the broader-ranging demands of fully conscious perceptual discrimination.
But whilst opponent-processing is another extraordinarily complex undertaking, even together, trichromatic and opponent-processing fall far short of supporting the plethora of deductions, inferences and anticipatory functions that underpin human visual sensibilities.
So before attempting a brief description of opponent-processing, let’s be clear about what lies ahead. To produce the everyday experience of the visual world that enables human beings to undertake yet more complex sense-making tasks, the stream of colour information considered so far needs substantial embellishment. The forms of colour information assembled within the retina will need to:
- Feed into and support the entire range of distinct attributes of visual perception and be available to be resolved within the composition of our immediate present visual reality as and when needed.
- Correspond with our extraordinary discriminatory capacities that interrogate a palette that can include millions of potentially observable variations in colour.
- Provide colour information about the outside world that is highly detailed, specifically relevant, spatially and temporally sequenced and capable of being cross-referenced with higher mental functions.
- Be subjectively orientated in a way that provides the impeccably accurate detail needed to deal with our more insane intentions. Think for example of leaping from rock to rock across a raging waterfall or diving off a mountain-top wearing only a wingsuit and negotiating the topography of craggy landscapes speeding past at 260km per hour.
Within the context of such extremely broad demands, much more is needed than the efficient transmission of the cleanly encode colour information that trichromatic phototransduction provides. The next stage involves the addition of contextualising detail and off more spacial and temporal information relevant to everyday circumstances. And as well as all this, the output must be capable of being handled within the data-carrying capacity of the optic nerves embedded in the back of each eye.
Opponent-processing of colour involves a sort of either/or interrogation of the infinitesimally small electrical impulses received from the three-channel output of the trichromatic investigation undertaken by bipolar calls and involves a biological equivalent of algorithms. Put simply, each signal received by a ganglion cell must pass three simultaneous tests that go something like this:
- Are you carrying information about the presence of more or less light than at other locations in your immediate area?
- Are you carrying information about the presence of a kind of red-green colour-contrast than at other locations in your immediate area?
- Are you carrying information about the presence of a kind of blue-yellow colour-contrast than at other locations in your immediate area?
Now, remember that the three cone types respond to a range of wavelengths within different bands of the visual spectrum with peak sensitivity centred on a very narrow range of wavelengths. With this in mind, it is reasonably easy to visualise that the incoming trichromatic traffic can be represented as a plot on a graph showing the intensity of response at every wavelength being surveyed by each cone type. If the original stimulus was produced by sunlight, containing wavelengths from right across the spectrum, then the graph for each cone type will show a curve across its range of sensitivity with a spike at peak sensitivities of L, M and S cones.
It is also clear that the simultaneous interrogation by ganglion cells of all three curves, and by cross-referencing each impression at multiple scales across the entire encoded retinal image, causes a quantity and quality of useful data to emerge that puts even the most sophisticated camera to shame. How? Because as this process proceeds in parallel with the broader discriminatory processes at work, every bit of colour information is already being invested with rudimentary forms of meaning!
Once the required level of organisation has been achieved, the encoded information exits the retina and is channelled along its respective optic nerve.
The two optic nerves contain around a million nerve fibres each and independently transmit composited information about every location within the visual field of their respective eye. At subsequent processing centres, all these separate packets of information will somehow be assembled into an image, with each signal in its proper spatial and temporal location, so that we see a stereoscopic picture of the world, updated many times a second. More than this, our impressions will seamlessly include information about the sound, feel, smell or tastes that relate to each matter of interest.
Imagine the amount of processing required to build coherent and fault-free images of our surroundings at this level of detail, all resolved into a continuous movie-like stream packed with meaning precisely relevant to our immediate needs! Imagine what it means to maintain this level of intelligence-saturated acuity, every day through the entire course of our lives!
Don’t forget, that as all this is going on, the images we will see are constantly being stabilized as we move our heads from side to side. Again, as our eyes scan left and right or up and down in their sockets, a single impression is sustained. And at any moment we may blink, focus on something else, or recompose everything we see before us as priorities change.
Let’s map some of these ideas out as bullet points.
-
- The description of human vision as a trichromatic colour system or as an opponent-process colour system refers to two complementary theories that help to explain aspects of colour perception that occur in the retina.
- Our understanding of trichromatic and opponent processes are called theories because our human understanding of exactly what happens at these infinitesimally small cellular scales can only be fully observed empirically and scientific enquiry is the circuitry is ongoing.
- There are four different photosensitive receptors commonly associated with the human retina, three types of cone cells and one type of rod cell. Cones are responsible for the perception of colour whilst rods function mainly in dim light.
- Cones work at their best at high levels of illumination.
- When lighting is low, rods come into their own. Because the rods are more sensitive to short wavelengths than to long ones, rods respond to the blue end of the visible spectrum. At night the scenes we see are registered as being relatively colourless.
- Too much light causes rods to saturate. Saturation in this context means that once the amount of light reaches a certain level the rods max-out and fail to register light levels rising further. This is where the cones take over.
- A distinction can be made between types of vision that result from adaption to light levels by the photosensitive rod and cone receptors:
- Photopic vision: This is where cone cells are most active. It refers to adaptation to higher levels of overall brightness within the field of view that an observer has adjusted to.
- Scotopic vision: This is where the rod cells are most active. It refers to adaptation to low levels of overall brightness within the field of view that an observer has adjusted to. Under these conditions, the ability to discern colours is sharply reduced. In addition, since there are no rods in the fovea and the cones there are not receiving enough light to be stimulated, the ability to discern fine detail is also compromised.
- Mesopic vision: This term refers to a combination of photopic vision and scotopic vision during which both rods and cones are active. There is no sharp transition at the top and bottom of the range of mesopic vision.
- Adaption refers to the fact that it can take as long as 20-30 minutes for our eyes to fully adjust to new light levels. This is reflected in the way we squint when going out into bright sunshine and by the fact that we may initially see little or nothing when indoor lights are turned out at night.
- Each of the millions of cone cells in the human eye can at any moment respond separately as photons strike them at light-speed. This is why a vast array of different colours can be perceived simultaneously at a single glance.
- Colour vision depends on bipolar and ganglion cells receiving input from at least two types of cone cells to produce trichromatic vision.
- People who have only two types of cone cells have dichromatic vision and are able to distinguish thousands rather than millions of different colours.
1.6 - Mixing light & pigments
Mixing light & pigments
As explained above, trichromatic processing of light entering the eye involves three types of cone receptors in the retina working in concert with bipolar cells to carry out an initial stage of colour processing.
One of the more important implications of trichromacy is that it is possible to match almost any naturally perceived colour by projecting wavelengths of light corresponding with red, green and blue onto a screen or surface so that the resulting admixture is reflected into the eyes.
We are exposed to mixtures of wavelengths of light corresponding with red, green and blue whenever we look at a phone or computer or any other type of digital screen. As the intensities of each of these three wavelengths are increased or decreased we see different colours. It is an astonishing fact that simply by manipulating different combinations, some digital screens are able to display a total of 16.78 million colours by combining 256 settings for red with 256 for green x 256 for blue.
A similar process takes place when we look at magazines or other forms of digital printing. In this case, cyan, magenta and yellow inks are used in place of light and different colours are observed as the amount of each pigment is adjusted. Digital printing also uses at least one additional black ink to add tonal depth to printed colours. Digital printing usually involves applying tiny dots of colour onto white paper so that the colours overlay one another and partially or entirely prevent incident light from being reflected off the surface and into our eyes.
There is an important difference between using red, green and blue light sources to produce colour on digital screens and using cyan, magenta and yellow inks for printing. When RGB light sources are added together in different proportions they cause our eyes to produce the impression of all the colours in the visible spectrum. When inks overlay one another they subtract from the range of possible wavelengths that can reflect off the paper. When cyan and magenta are overlayed they reduce the amount of blue light that reaches our eyes, when cyan and yellow are overlayed they reduce the amount of green light that reaches our eyes, and when magenta and yellow are overlayed they reduce the amount of red that reaches our eyes. When CMY are overprinted they prevent red, green and blue from reaching our eyes.
- A human observer with trichromatic vision exposed to a mixture of the three wavelengths of light corresponding with red, green and blue can see a full range of colours if they are combined in the correct proportions. Red, green and blue, when used in this way, are called additive primary colours. This is the basis of the RGB colour model.
- A human observer with trichromatic vision exposed to a mixture of three pigments corresponding with cyan, magenta and yellow can see a full range of colours if they are mixed together in the correct proportions. Cyan, magenta and yellow, when used in this way, are called subtractive primary colours. This is the basis of the CMY colour model.
- All vision-related technologies take advantage of the trichromatic nature of colour vision and of the remarkable discoveries that are associated with it.
- The screens of televisions, computers, mobile phones and digital projectors all mix wavelengths of light corresponding with red, green and blue (RGB), to produce the richly coloured images that we take so much for granted.
- Digital printing meanwhile uses cyan, magenta and yellow inks (CMY) inks but also adds black (CMYK) to produce the appearance of deeper shadows on the printed page.
- Additive colour mixing is not only applied to the output of colour information to digital screens and paper. Sensors in cameras and scanners work in a similar way, encoding and recording inputs on red, green and blue channels.
The terms trichromacy and trichromatic processing are used to describe the biological operations taking place within our eyes. When this underpinning of colour vision is exploited to intentionally match colours we want to mix and display it is often referred to as a tristimulus system of colour matching. Clearly, both the RGB and CMY colour models are tristimulus systems.
- A tristimulus system visually matchs a colour perceived under standardized conditions against mixtures of the three primary colours belonging to a chosen colour model.
- Each colour model has methods of notation for keeping track of the exact amounts of the primary colours needed to match a perceived colour.
1.7 - Additive and subtractive colour
- Whilst additive colour mixing is the method used to combine wavelengths of light, subtractive colour mixing is the method used with dyes, inks and pigments.
- An additive approach to colour mixing is used in the case of the emission of light by light-emitting diodes (or similar light sources) embedded into the screens of mobile phones, computers and televisions etc.
- An additive approach to colour mixing is also used with digital projectors. In this case, sufficient light must be produced on each channel to form intense images when focused onto a screen across a room.
- RGB colour is one of the additive colour models that combine wavelengths of light corresponding with red, green and blue primary colours to produce other colours.
- Red, green and blue are called additive primary colours in an RGB colour model because they can be added together to produce other colours. Red green and blue are often described as being components of the resulting colour.
- Different colours are produced by varying the intensity of the component colours between fully off and fully on.
- When fully saturated red, green and blue primary colours are combined, they produce white.
- When any two fully saturated additive primaries are combined, they produce a secondary colour: yellow, cyan and magenta.
- Some RGB colour models can produce over 16 million colours by fine-tuning the intensity of each of the three primary colours.
- The additive RGB colour model cannot be used for mixing different colours of pigments, paints, inks, dyes or powders. To combine these colourants subtractive colour models are used.
1.8 - Trichromacy and tristimulus systems
Trichromatic colour model
Trichromatic colour models (and the trichromatic colour theory that underpins them) provide methods for visually matching and mixing colours from combinations of three primary colours – red, green and blue ( or cyan, magenta and yellow). The information about how much of each primary colour is needed to produce a target colour is stored as tristimulus values. Tristimulus values are simply codes that can be used to record and pass on colour information.
The LMS colour model (long, medium, short), is a trichromatic colour model that represents the response of the three types of cones of the human eye, named for their responsivity (sensitivity) peaks at long, medium, and short wavelengths. It is used to systematize the response of the three types of cones of the human eye to different visual stimuli, that is, different wavelengths of light. The strength of the LMS colour model is its concern for the connection between the physiological aspects of vision and the everyday visual experience of an observer. L M and S refer to the bands of wavelengths that each cone type within the retina responds to.
Let’s look more carefully at this connection between trichromacy and tristimulus systems of which the RGB colour model provides a good example.
We start with the premise that trichromatic processing within the retina reduces all colours an observer sees to responses corresponding with the spectral biases of L, M and S cone cells.
This premise can be demonstrated experimentally by positioning an observer in front of three different light torches, each covered with a red, green or blue filter, that project light onto the same area of a neutrally coloured surface. The effect of each filter is to block all wavelengths except one. If the three torches project light at equal intensities the surface appears white. If the intensity of light or the colours of the filters are not exactly matched a colour cast will be apparent. If one light is turned off, then a secondary colour appears. Depending on which colour is absent the result will be cyan, yellow or magenta.
The reason the surface appears white to the observer in this experiment when all three torches are turned on is that each of the three cone types in their retina is being triggered evenly. This means that each of these types of photosensitive neurons are registering the presence of the wavelengths of light they are tuned to.
In the second part of the demonstration, a calibrated dial is used to alter the intensity of each torch. By setting each dial to a component of a tristimulus value for a known colour, it is possible to test whether the resulting stimulus causes the observer to see the intended colour.
This experiment corresponds directly with the way all RGB devices such as TVs, computer monitors, phone screens and projectors work in so far as tristimulus RGB values are used to stimulate the L, M and S cone cells on the retina to produce the intended experience of colour.
Opponent processing does not play a determining role in this experiment. We know from opponent-processing theory that after trichromatic processing takes place, the signals will be processed based on whether the cone responses indicate that the stimulus is bright or dull, more red-or-green, and at the same time, more blue-or-yellow. The output of this process will be fed into the million-or-so fibres of each optic nerve encoded into two channels of chromatic information and one dealing with the perception of brightness.
Experiments by several generations of scientists and artists have confirmed the connection between trichromacy and tristimulus systems. Opponent-processing cannot be demonstrated quite so directly but visual illusions and unexpected consequences of different attributes of colour perception have been used experimentally to unravel what is going on with extraordinary success.
One of the outcomes of research into tristimulus systems is the requirement, when choosing primary colours, that two of them cannot be combined to produce the third. Each must be unique so far as the human eye is concerned.
Research into the opponent-process has established that there are in fact four unique colours, red, green, blue and yellow, each of which shows no perceptual similarity to any of the others.
The implications of the fact that human vision can be stimulated by three distinct colour inputs are:
- In normal conditions, any particular colour seen by an observer is produced by complex patterns of different wavelengths and intensities of light from across the visible spectrum as they enter the eye and are absorbed by cone cells within the retina in real-time.
- The complex pattern of wavelengths and intensities of light being emitted by a light source at any moment is called its spectral power distribution. A spectral power distribution can be plotted on a graph and always appears as a wavy line with red at one end and violet at the other. The profile of the plot rises for high and falls for low intensities of light.
- The colour notation used to record tristimulus values can, in principle, describe any human colour sensation.
- If tristimulus values corresponding with the full range of human observable colours are plotted on a graph, with three axes drawn perpendicular to one another, they can produce an inclusive representation of colour perception in the form of a 3-dimensional colour solid.
- The three axes correspond with the range of responses of the three cone types and so can be labelled L, M and S. A scale along each axis can be added to correspond with a minimum cone response at one end and a maximum at the other. This is the basis of the LMS colour model, which is one of a number of colour models devised to quantify human colour vision.
Colour models such as RGB colour and the Munsell colour system also use tristimulus notation to record colour information. The implications are that LMS, RGB and Munsell are all grounded in the trichromatic nature of human vision and take advantage of the resulting opportunities in terms of systems that use additive colour.
Other colour models such as HSB colour, HSV colour and HSL colour which are all variants of RGB colour do not use forms of notation that correspond directly with tristimulus value.
1.9 - LMS & RGB colour models
LMS & RGB colour models
The LMS colour model is used to represent the response of the three types of cones of the human eye to different visual stimuli, that is, different wavelengths of light.
The strength of the LMS colour model is its concern for the connection between the physiological aspects of vision and the everyday visual experience of an observer.
LMS (long, medium, and short) refers to the band of wavelengths that each cone type within the retina responds to. The LMS model is particularly useful when comparing an observer’s response to the appearance of colour samples.
The RGB colour model and derivatives such as the HSB colour model, build on the knowledge gained from research into the response of the human eye to trichromatic stimuli and so has foundations in LMS’s concern with the biological processing of colours within the human eye.
Design and imaging software such as Adobe Creative Colour rarely refer to LMS and doesn’t include LMS-related tools but it underpins colour models such as the RGB, HSB and CMY colour models.
The functionality of the RGB colour model is rooted in the parallel notion that information about any colour whatsoever can be described in terms of the amount of red, green and blue it contains.
When using decimal notation to record the amounts of R, G and B needed to produce any observable colour, R, G and B are each assigned a value between 0 and 255. In the case of the output of this information to a computer screen, each value is sent along a separate low-voltage electrical circuit with a direct correspondence between the intended colour value and amperage ( low amps start at zero, high amps max at 255). As the amps change on each channel so does the brightness of a red, green or blue light-emitting diode (LED’s) embedded in each circuit.
Digital screens work on the basis of sets of three diodes grouped into pixels embedded into the screen. A typical 4K screen might contain 2160 x 3840 pixels so a total of 8.29 million. The colour of each pixel is hardwired into the technology so the wavelength of light emitted by each diode is fixed. It is therefore simply the relationship between how dark a screen appears when pixels are off and the fine control over the intensity of light they are able to produce that determines the range of colours that can be produced in the eyes of the beholder. The total range of colours is often referred to as a gamut.
Different styles of notation are used to record, store and transmit calibrated colour information depending on the type and purpose of the colour model in use. Adobe Creative Cloud software, for example, used one system for RGB values and another for HSB.
RGB colour values are typically represented by decimal triplets (base 10) or hexadecimal triplets (base 16) and always ordered red, then green, then blue.
Decimal numbers from 0 to 255 are selected for each value. Zero means fully off and 255 means maximum intensity:
- R=255, G=00, B=00 or 255,00,00: Red
- R=255, G=255, B=00 or 255,255,00: Yellow
- R=00, G=255, B=00 or 00, 255,00: Green
- R=00, G=255, B=255 or 00,255,255: Cyan
- R=00, G=00, B=255 or 00,00,255: Blue
- R=255, G=00, B=255: 255,00,,255: Magenta
In hexadecimal notation, the same triplets appear as follows. Hexadecimal numbers range from 00 to FF:
- #FF 00 00: Red
- #FF FF 00: Yellow
- #00 FF 00: Green
- #00 FF FF: Cyan
- #00 00 FF: Blue
- #FF 00 FF: Magenta
The hash symbol (#) indicates hex notation.
The sequence of hexadecimal values between 0 and 15 = 0,1, 2, 3, 4, 5, 6, 7, 8, 9, A, B, C, D, E and F.
The next sequence of hexadecimal values between 16 and 31 = 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 1A, 1B, 1C, 1D, 1E and 1F.
As a footnote, it is worth recalling that as we sit engrossed in the unfolding narrative of a movie on a digital display the flow of images is produced by a matrix of pixels each of which is driven by a succession of tristimulus values that typically refresh 240 times a second. 8.29 million pixels updating 240 times per second means we are actually watching around 250 million flickering colours every second. This moving image is then focused by the lens in each eye and produces a perfect miniature replica on the retinal surface of each eye. As the 6 million photosensitive cone cells per eye react and phototransduction takes place When an observer looks at the screen, the image is focused on the retina at the back of the eyeball by the lens to enable the S, M, and L cone cell types to capture as much information as possible. As we know, cones are also organised into the biological equivalent of a matrix so that responses to each pixel within the image that has been projected onto the retina can be accurately mapped and each pixel-point of light arranged in its proper place within the living experience of the observer.
It is the challenges of finding ways to determine the tristimulus values needed to produce the appropriate stimuli to enable an observer to see or choose a particular colour, or range of colours, in different viewing conditions that have resulted in the development of competing colour systems each with its own colour model and associated colour spaces. The equipment and devices we use every day are critically judged on the quality of the images they produce.
It is these same challenges that have led to the development of colour profiles that ensure, for example, that the relationship between the original colours seen by an observer taking a photograph and colours that appear after processing are reproduced as intended. Colour profiles ensure that as information is encoded and passes from device to device, every colour within a range of colours (a gamut) appears as expected when output by a printer regardless of its make or model.
It is worth noting that there is no standard set of primary light sources or pigments sufficient to enable an observer to see all colours within the gamut of human vision in every situation. As a result, the choice of light sources and pigments is adjusted to suit different technologies, media and viewing conditions.
https://en.wikipedia.org/wiki/RGB_colour_model
http://www.huevaluechroma.com/014.php
https://en.wikipedia.org/wiki/Trichromacy
https://en.wikipedia.org/wiki/Colour_vision
ANATOMY OF THE EYE
2.1 - Retina
THE RETINA
Human beings see the world in colour because of the way their visual system processes light. The retina contains light-sensitive receptors, rod and cone cells, that respond to light stimuli. It is the variety of wavelengths and intensities of light entering the eyes that produces the impression of colour.
The retina is the innermost, light-sensitive layer of tissue inside our eyes. It forms a sheet of tissue barely 200 micrometres (μm) thick, but its neural networks carry out almost unimaginably complicated feats of image processing.
The physiology of the eye results in a tiny, focused, two-dimensional image of the visual world being projected onto the retina’s surface. Because of the optics of lenses, it appears upside down and the wrong way around. But no worry, sorting that out is child’s play for the human brain! The real challenge is that the photosensitive receptors in the retina must produce precise chemical responses to light and translate every minute detail of the image into electrical impulse ready to be sent to the brain where they produce visual impressions of the world. In a very limited sense, the retina serves a similar function to a photosensitive chip in a camera.
As research continues to reveal ever-increasing amounts of detail about these signalling processes across and beyond the retina, it required new thinking, not only of the retina’s function but also of the mechanisms within the brain that shape these signals into behaviourally useful perceptions.
The retina consists of 60-plus distinct neuron types, each of which plays a specialized role in turning variations in the patterns of wavelengths and intensities of light into visual information. Neurons are electrically excitable nerve cells that collect, process and transmit vast amounts of this information through both chemical and electrical signals. Retinal neurons work together to convert the signals produced by a hundred and twenty million rods and cones and send them along around one million fibres within the optic nerve of each eye to connections with higher brain functions. In this process rods and cones are first responders whilst ganglion cells are the final port of call before information leaves the retina.
There are three principal forms of processing that take place within the retina itself. The first organises the outputs of the rod and cone photoreceptors and begins to compose them into around 12 parallel information streams as they travel through bipolar cells. The second connects these streams to specific types of retinal ganglion cells. The third modulates the information using feedback from horizontal and amacrine cells to create the diverse encodings of the visual world that the retina transmits towards the brain.
As mentioned above, the image of the outside world focused on the retina is upside down and the wrong way around. But the human retina is also inverted in the sense that the light-sensitive rod and cone cells are not located on the surface where the image forms, but instead are embedded inside, where the retina attaches to the fabric of the eyeball. As a result, light striking the retina, passes through layers of other neurons (ganglion, bipolar cells etc.) and blood-carrying capillaries, before reaching the photoreceptors.
The overlying neural fibres do not significantly degrade vision in the inverted retina. The neurons are relatively transparent and accompanying Müller cells act as fibre-optic channels to transport photons directly to photoreceptors. However, some estimates suggest that overall, around 15% of all the light entering the eye is lost en-route to the retina. To counter this, the fovea centralis, at the centre of our field of vision, is free of rods and there are no blood vessels running through it, so optimising the level of detail where we need it most.
https://en.wikipedia.org/wiki/Retina
2.2 - Retinal input
Retinal input
Visual input is initially encoded in the retina as a two-dimensional distribution of light intensity, expressed as a function of position, wavelength and time. This retinal image is transferred to the visual cortex where primary sensory cues and, later, inferred attributes, are eventually computed in the process of actualising our perceptions. Parallel processing strategies are employed from the outset to overcome the constraints of the individual ganglion cell’s limited bandwidth and anatomical bottlenecks as data approaches the optic nerves that connect each eye to the visual cortex.
References: DeYoe and Van Essen (1988): https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2771435/
2.3 - Rods and cones
Rods and cones
Both the photosensitive rods and cones form a regularly spaced mosaic of cells across the entirety of the retina – bar the absence of rods in the fovea centralis. Because there are 100 million rod receptors and 20 million cone receptors in each eye, rods are packed more densely per unit area. The synaptic connections of both rods and cones vary in function in different locations across the retina, reflecting the specialisations of different regions. This, for example, allows the eyes to deal with daylight and darkness and with what we see at the centre and periphery of our field of view.
Rods and cones are easily distinguished by their shape, from which they derive their names, the type of photo pigment they contain and by the distinct patterns of synaptic connections with the other neurons around them.
Neurons (nerve cells) are present throughout the human central and peripheral nervous systems and fall into three main categories: sensory, motor and interneurons. Rods and cones are both sensory neurons. Rods don’t produce as sharp an image as cone cells because they share more connections with other types of neurons. But a rod cell is believed to be sensitive enough to respond to a single photon of light whilst cone cells require tens to hundreds of photons to be activated.
The principal task of rod and cone cells alike is photo-transduction. This refers to the type of sensory transduction that takes place in the visual system. It is the process of photo-transduction that enables pigmented chemicals in the rods and cones to sense light and convert it into electrical signals. Many other types of sensory transduction occur elsewhere within the body enabling touch and hearing for example.
References: Functional Specialization of the Rod and Cone Systems: https://www.ncbi.nlm.nih.gov/books/NBK10850/
2.4 - Trichromatic colour vision
Trichromatic processing
To get some sense of what is going on during the trichromatic processing of visual information collected during phototransduction within rod and cone cells, we need to imagine the light-receiving layer within the retina of each eye as being composed of around 120 million points. Each point is the location of a rod or cone within a mosaic of cells stretching to the boundary of the retina in every direction. Each point also corresponds with a different location within the overall image of the world outside that is projected onto the retina by the lens as light enters the eyes through the cornea, and pupil.
Every rod and cone cell within the human eye is an independent photo-sensitive neuron that responds independently to incoming photons of light. In the first instance, each one carries out its task of responding to the stream of different wavelengths of light without reference to the receptors around it. We must imagine all these neurons as being capable of functioning simultaneously and being able to fire several times a second. Each time a cell responds, phototransduction takes place, which is a chemical response to light that produces electrical impulses ready for transmission.
What precisely happens depends on lighting conditions.
When light levels are low, each photosensitive rod cell may be hit by a handful of photons of a given wavelength and in some cases, this is enough to trigger the chemical response. But at very low levels of lighting, the response of cone cells is very limited. The outcome is a typical night scene that appears blue-purple-black with very little detail.
When light levels are high, each cone cell may be bombarded with tens of thousands of photons of a given wavelength within the briefest fraction of a second, producing a powerful chemical response. In these conditions, rod cells tend to be overwhelmed by the sheer quantity of light but can still provide useful information in peripheral vision. The outcome is a world rich in colour, full of detail and with contrasting highlight-detail and deep well-defined areas of shadow.
The trichromatic process involves the sampling of this mosaic of rod and cone photoreceptors by bipolar and horizontal cells. Individual photoreceptors are connected in small groups to bipolar cells that receive their electrical output. The bipolar cells compare the response of receptors with the response of neighbours, whilst horizontal cells help to aggregate the result and encode it into three independent data channels.
The comparison process involves what is termed centre-surround antagonism. This refers to the way bipolar neurons organize their receptive fields. A close-up view reveals that centre-surround antagonism relies on small groups of cells being arranged around a centre point where one rod or cone synapses on the dendrites of a bipolar cell. Around this centre are other photoreceptors that also synapse onto other dendrites of the same bipolar cell. The signal received from the centre is compared with a summation of the signals from neighbours to establish to what extent they agree or disagree. This process goes on in real-time as bipolar cells receive successions of signals and horizontal cells modulate the information to improve fidelity. The scale of this enterprise as it takes place across the surfaces of the retina in each eye and in real-time is extraordinary.
The role of horizontal cells in both trichromatic and centre-surround antagonism is best conceived as signal conditioning and particularly with globally adjusting visual information ready for opponent processing. However, ongoing research suggests that horizontal cells may also be involved in an early stage of signal interpretation and so contribute towards things such as the detection of edges and movement.
Trichromatic colour vision is governed by the fact that there are three distinct types of cone cells within the retina, each tuned to respond to a different band of wavelengths of light. As light floods in, each type of cone outputs a signal if it picks up the presence of photons with wavelengths it recognises. Every identifiable point on the retina contains all three types of cone cells and these are tightly packed into a random mosaic pattern across the entire surface. The result is a photosensitive film containing millions of receptors capable of responding to all wavelengths across the visible spectrum.
General descriptions of trichromatic vision often suggest that the three types of cone cells in the human retina are responsive to wavelengths corresponding to red, green and blue. It is more accurate to say that the peak sensitivity of these L, M and S cones types respond with biases towards different regions of the visible spectrum and have a loose correspondence with red, green and blue:
- L cones: Respond to long wavelengths (peak sensitivity around 560 – 580 nanometres) so with a region of sensitivity that includes red, orange, green and yellow but with a peak bias between red and yellow.
- M cones: Respond to medium wavelengths (peak sensitivity around 530 – 545 nanometres) so with a region of sensitivity that includes orange, green, yellow and cyan but with a peak bias between yellow and green.
- S cones: Respond to short wavelengths (peak sensitivity around 420 – 440 nanometres) so with a region of sensitivity that includes cyan, blue and violet but with a peak bias between blue and violet.
- Rods: Rod cells are most sensitive to wavelengths around 498 nanometres, so with a peak sensitivity towards green-blue, and are insensitive to wavelengths longer than about 640 nanometres (red).
This arrangement suggests that:
- At any specific moment, all three types of cone cells at any specific location on the retina may fire multiple times per second in response to streams of photons that have constantly changing wavelengths.
- The assessment of the distribution of wavelengths by every individual cone depends on both its range of sensitivity and the wavelength at which its response peaks.
- Centre-surround comparisons check for consistencies and variations within the responses of cone groupings at every location.
- As soon as a clean and noise-free consensus is achieved, the data specific to each cone type is output on one of three separate data channels.
- Once the composite of trichromatic and centre-surround encoding of colour information is complete it is sent onward for opponent processing within the retina.
- Recent research suggests that the three output channels do not correspond directly with the L, M and S cone types. There are indications that two channels contain chromatic information whilst the other contains achromatic data.
2.5 - Trivariance
Trivariance
The term trivariance is used to refer to this first stage of the trichromatic process. It refers to both the phototransductive response of the cone cells themselves and to the three separate channels used to convey their colour information forward to subsequent levels of neural processing.
Each channel conveys information about the response of one cone-type to both the wavelength of the incoming light it is tuned to and to its intensity. In both physiological and neurological terms this process is exclusively concerned with trivariance – three discernible differences in the overall composition of light entering the eye.
It is the separation of the signals produced on each channel that accounts for the ability of our eyes to respond to stimuli produced by additive mixtures of wavelengths corresponding with red, green and blue primary colours. But more of that later!
By way of summary, the rod and trivariant cone systems are composed of photoreceptors with connections to other cell types within the retina. Both specialize in different aspects of vision. The rod system is extremely sensitive to light but has a low spatial resolution. Conversely, the cone system is designed to function in stronger light. As a result, cones are relatively insensitive compared with rods but have a very high spatial resolution. It is this specialisation that results in the extraordinary detail, resolution and clarity of human vision.
| Rod System | Cone System |
| High sensitivity, specialized for night vision | Lower sensitivity specialized for day vision |
| Saturate in daylight | Saturate only in intense light |
| Achromatic | Chromatic, mediate colour vision |
| Low acuity | High acuity |
| Not present in the central fovea | Concentrated in the central fovea |
| Present in larger number than cones | Present in smaller number than rods |
Caption
2.6 - Retinal image
Retinal image
It is the cornea-lens system that determines where light falls on the surface of the retina which results in discernible images.
The images are inverted and obviously very small compared with the world outside that they resolve. The inversion poses no problem. Our brains are very flexible and even when tricked by prisms will always turn the world right-side-up given time. The reduction in size is part of the process by which the fit of the image on the retina determines our field of view.
The images are real in the sense that they are formed by the actual convergence of light rays onto the curved plane of the retina. Only real images of this kind provide the necessary stimulation of rod and cone cells necessary for human perception.
2.7 - Fovea centralis
Fovea centralis
The entire surface of the retina contains nerve cells, but there is a small portion with a diameter of approximately 0.25 mm at the centre of the macula called the fovea centralis where the concentration of cones is greatest. This region is the optimal location for the formation of image detail. The eyes constantly rotate in their sockets to focus images of objects of interest as precisely as possible at this location.
2.8 - Accommodation
Accommodation
The distance between the retina (the detector) and the cornea (the refractor) is fixed in the human eyeball. The eye must be able to alter the focal length of the lens in order to accurately focus images of both nearby and far away objects on the retinal surface. This is achieved by small muscles that alter the shape of the lens. The distance of objects of interest to an observer varies from infinity to next to nothing but the image distance remains constant.
The ability of the eye to adjust its focal length is known as accommodation. The eye accommodates by assuming a lens shape that has a shorter focal length for nearby objects in which case the ciliary muscles squeeze the lens into a more convex shape. For distant objects, the ciliary muscles relax, and the lens adopts a flatter form with a longer focal length.
CELULAR COMPOSITION
3.1 - Bipolar cells
Bipolar cells
Bipolar cells, a type of neuron found in the retina of the human eye connect with other types of nerve cells via synapses. They act, directly or indirectly, as conduits through which to transmit signals from photoreceptors (rods and cones) to ganglion cells.
There are around 12 types of bipolar cells and each one functions as an integrating centre for a different parsing of information extracted from the photoreceptors. So, each type transmits a different analysis and interpretation of the information it has gathered.
The output of bipolar cells onto ganglion cells includes both the direct response of the bipolar cell to signals derived from photo-transduction but also responses to those signals received indirectly from information provided by nearby amacrine cells that are also wired into the circuitry.
We might imagine one type of bipolar cell connecting directly from a cone to a ganglion cell that simply compares signals based on differences in wavelength. The ganglion cell might then use the information to determine whether a certain point is a scene is red or green.
Not all bipolar cells synapse directly with a single ganglion cell. Some channel information that is sampled by different sets of ganglion cells. Others terminate elsewhere within the complex lattices of interconnections within the retina so enabling them to carry packets of information to an array of different locations and cell types.
3.2 - Amacrine cells
Amacrine cells
Amacrine cells interact with bipolar cells and/or ganglion cells. They are a type of interneuron that monitor and augment the stream of data through bipolar cells and also control and refine the response of ganglion cells and their subtypes.
Amacrine cells are in a central but inaccessible region of the retinal circuitry. Most are without tale-like axons. Whilst they clearly have multiple connections to other neurons around them, their precise inputs and outputs are difficult to trace. They are driven by and send feedback to the bipolar cells but also synapse on ganglion cells, and with each other.
Amacrine cells are known to serve narrowly task-specific visual functions including:
- Efficient transmission of high-fidelity visual information with a good signal-to-noise ratio.
- Maintaining the circadian rhythm, so keeping our lives tuned to the cycles of day and night and helping to govern our lives throughout the year.
- Measuring the difference between the response of specific photoreceptors compared with surrounding cells (centre-surround antagonism) which enables edge detection and contrast enhancement.
- Object motion detection which provides an ability to distinguish between the true motion of an object across the field of view and the motion of our eyes.
Centre-surround antagonism refers to the way retinal neurons organize their receptive fields. The centre component is primed to measure the sum-total of signals received from a small number of cones directly connected to a bipolar cell. The surround component is primed to measure the sum of signals received from a much larger number of cones around the centre point. The two signals are then compared to find the degree to which they agree or disagree.
3.3 - Horizontal cells
Horizontal cells
Horizontal cells are connected to rod and cone cells by synapses and are classed as laterally interconnecting neurons.
Horizontal cells help to integrate and regulate information received from photoreceptor cells, cleaning up and globally adjusting signals passing through bipolar cells towards the regions containing ganglion cells.
An important function of horizontal cells is enabling the eye to adjust to both bright and dim light conditions. They achieve this by providing feedback to rod and cone photoreceptors about the average level of illumination falling onto specific regions of the retina.
If a scene contains objects that are much brighter than others, then horizontal cells are believed to prevent signals representing the brightest objects from dazzling the retina and degrading the overall quality of information.
Caption
The Neuronal Organization of the Retina Richard H. Masland
https://www.cell.com/neuron/fulltext/S0896-6273(12)00883-5?_returnURL=https%3A%2F%2Flinkinghub.elsevier.com%2Fretrieve%2Fpii%2FS0896627312008835%3Fshowall%3Dtrue
3.4 - Ganglion cells
Ganglion cells
Retinal ganglion cells are located near the boundary between the retina and the central chamber containing vitreous humour. They collect and process all the visual information gathered directly or indirectly from the forty-something types of rod, cone, bipolar, horizontal and amacrine cells and, once finished, transmit it via their axons towards higher visual centres within the brain.
The axons of ganglion cells form into the fibres of the optic nerve that synapse at the other end on the lateral geniculate nucleus. Axons take the form of long slender fibre-like projections of the cell body and typically conduct electrical impulses, often called action potentials, away from a neuron.
A single ganglion cell communicates with as few as five photoreceptors in the fovea at the centre of the macula. This produces images containing the maximum possible resolution of detail. At the extreme periphery of the retina, a single ganglion cell receives information from many thousands of photoreceptors.
Around twenty distinguishable functional types of ganglion cells resolve the information received from 120 million rods and cones into one million parallel streams of information about the world surveyed by a human observer in real-time throughout every day of their lives. They function to complete the construction of the foundations of visual experience by the retina, ordering the eyes response to light into the fundamental building blocks of vision. Ganglion cells do the groundwork that enables retinal encodings to ultimately converge into a unified representation of the visual world.
Ganglion cells not only deal with colour information streaming in from rod and cone cells but also with the deductions, inferences, anticipatory functions and modifications suggested by bipolar, amacrine and horizontal cells. Their challenge, therefore, is to enable all this data to converge and to assemble it into high fidelity, redundancy-free, compressed and coded form that can continue to be handled within the available bandwidth and so the data-carrying capacity of the optic nerve.
It is not hard to imagine the kind of challenges they must deal with:
- Information must feed into and support the distinct attributes of visual perception and be available to be resolved within the composition of our immediately present visual impressions whenever needed.
- Information must correspond with the outstanding discriminatory capacities that enable the visual system to operate a palette that can include millions of perceivable variations in colour.
- Information about the outside world must be able to be automatically cross-referenced, highly detailed, specifically relevant, spatial and temporally sequenced and available on demand.
- Information must be subjectively orientated in a way that it is locked at an impeccable level of accurate detail to even our most insane intentions as we leap from rock to rock across a swollen river or dive from an aircraft wearing only a wingsuit and negotiate the topography of a mountainous landscape speeding past at 260km per hour.
It is now known that efficient transmission of colour information is achieved by a transformation of the initial three trivariant colour mechanisms of rods and cones into one achromatic and two chromatic channels. But another processing stage has now been recognised that dynamically readjusts the eye’s trivariant responses to meet criteria of efficient colour information management and to provide us with all the necessary contextualising details as we survey the world around us. Discussion of opponent-processing is dealt with in the next article!
3.5 - Müller cells
Müller cells
Müller glia, or Müller cells, are a type of retinal cell that serve as support cells for neurons, as other types of glial cells do.
An important role of Müller cells is to funnel light to the rod and cone photoreceptors from the outer surface of the retina to where the photoreceptors are located.
Other functions include maintaining the structural and functional stability of retinal cells. They regulate the extracellular environment, remove debris, provide electrical insulation of the photoreceptors and other neurons, and mechanical support for the fabric of the retina.
- All glial cells (or simply glia), are non-neuronal cells in the central nervous system (brain and spinal cord) and the peripheral nervous system.
- Müller cells are the most common type of glial cell found in the retina. While their cell bodies are located in the inner nuclear layer of the retina, they span the entire retina.
3.6 - Pigment epithelium
Pigment epithelium
Pigment epithelium is a layer of cells at the boundary between the retina and the eyeball that nourish neurons within the retina. It is firmly attached to the underlying choroid is the connective tissue that forms the eyeball on one side but less firmly connected to retinal visual cells on the other.
OPTICAL PATHWAY
4.1 - Optic nerve
Optic nerve
The optic nerve is the cable–like grouping of nerve fibres formed from the axons of ganglion cells that transmit visual information towards the lateral geniculate nucleus.
The optic nerve contains around a million fibres and transports the continuous stream of data that arrives from rods, cones and interneurons (bipolar, amacrine cells). The optic nerve is a parallel communication cable that enables every fibre to represent distinct information about the presence of light in each region of the visual field.
4.2 - Optic chiasm
Optic chiasm
The optic chiasm is the part of the brain where the optic nerves partially cross. It is located at the bottom of the brain immediately below the hypothalamus.
The cross-over of optic nerve fibres at the optic chiasm allows the visual cortex to receive the same hemispheric visual field from both eyes. Superimposing and processing these monocular visual signals allows the visual cortex to generate binocular and stereoscopic vision.
So, the right visual cortex receives the temporal visual field of the left eye, and the nasal visual field of the right eye, which results in the right visual cortex producing a binocular image of the left hemispheric visual field. The net result of optic nerves crossing over at the optic chiasm is for the right cerebral hemisphere to sense and process left-hemispheric vision, and for the left cerebral hemisphere to sense and process right-hemispheric vision.
4.3 - Lateral geniculate nucleus
Lateral geniculate nucleus
The lateral geniculate nucleus is a relay centre on the visual pathway from the eyeball to the brain. It receives sensory input from the retina via the axons of ganglion cells.
The thalamus which houses the lateral geniculate nucleus is a small structure within the brain, located just above the brain stem between the cerebral cortex and the midbrain with extensive nerve connections to both.
The lateral geniculate nucleus is the central connection for the optic nerve to the occipital lobe of the brain, particularly the primary visual cortex.
Both the left and right hemispheres of the brain have a lateral geniculate nucleus.
There are three major cell types in the lateral geniculate nucleus which connect to three distinct types of ganglion cells:
- P ganglion cells send axons to the parvocellular layer of the lateral geniculate nucleus.
- M ganglion cells send axons to the magnocellular layer.
- K ganglion cells send axons to a koniocellular layer.
The lateral geniculate nucleus specialises in calculations based on the information it receives from both the eyes and from the brain. Calculations include resolving temporal and spatial correlations between different inputs. This means that things can be organised in terms of the sequence of events over time and the spatial relationship of things within the overall field of view.
Some of the correlations deal with signals received from one eye but not the other. Some deal with the left and right semi-fields of view captured by both eyes. As a result, they help to produce a three-dimensional representation of the field of view of an observer.
- The outputs of the lateral geniculate nucleus serve several functions. Some are directed towards the eyes, others are directed towards the brain.
- A signal is provided to control the vergence of the two eyes so they converge at the principal plane of interest in object-space at any particular moment.
- Computations within the lateral geniculate nucleus determine the position of every major element in object-space relative to the observer. The motion of the eyes enables a larger stereoscopic mapping of the visual field to be achieved.
- A tag is provided for each major element in the central field of view of object-space. The accumulated tags are attached to the features in the merged visual fields and are forwarded to the primary visual cortex.
- Another tag is provided for each major element in the visual field describing the velocity of the major elements based on changes in position over time. The velocity tags (particularly those associated with the peripheral field of view) are also used to determine the direction the organism is moving relative to object-space.
4.4 - Optic radiation
Optic radiation
The optic radiations are tracts formed from the axons of neurons located in the lateral geniculate nucleus and leading to areas within the primary visual cortex. There is an optic radiation on each side of the brain. They carry visual information through lower and upper divisions to their corresponding cerebral hemisphere.
VISUAL CORTEX
5.1 - Primary visual cortex
Primary visual cortex
The visual cortex of the brain is part of the cerebral cortex and processes visual information. It is in the occipital lobe at the back of the head.
Visual information coming from the eyes goes through the lateral geniculate nucleus within the thalamus and then continues towards the point where it enters the brain. The point where the visual cortex receives sensory inputs is also the point where there is a vast expansion in the number of neurons.
Both cerebral hemispheres contain a visual cortex. The visual cortex in the left hemisphere receives signals from the right visual field, and the visual cortex in the right hemisphere receives signals from the left visual field.
[Cerebral hemispheres, occipital lobes, primary visual cortex, optical radiations]