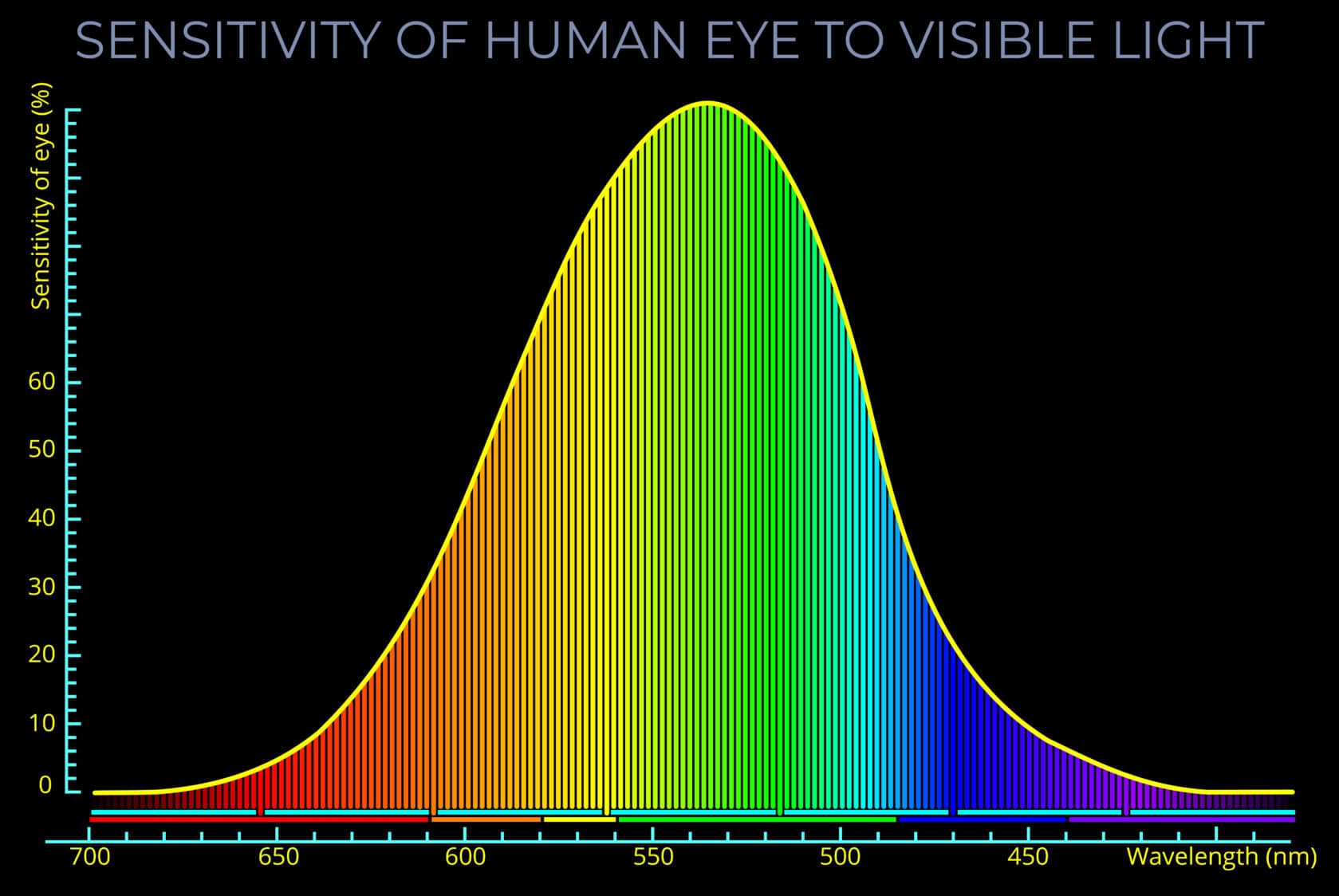
Vision – An introduction
$0.00
Here are all the diagrams in our Introduction to Vision series.
Each one appears on its own page with a full explanation.
AND
Did you know that all our diagrams are free to download!
Description
Vision - An introduction
Each diagram appears on its own page with a full explanation.
AND
Did you know all diagrams are available as FREE downloads?